
NF-κBは1986年に、David BaltimoreらによりB細胞で選択的に発現する免疫グロブリンのκ軽鎖のエンハンサーに結合する転写因子として同定されました。
NF-κBは免疫応答や細胞の生存など多彩な生命現象に関与しており、リウマチ・アレルギー疾患、がんなどでその活性が亢進していることから、2013年3月現在、Pubmedで50,000以上の論文が検索されるように、非常にextensiveに研究されている転写因子です。
NF-κBはRelファミリーに属するタンパク質のヘテロあるいはホモ2量体です。NF-κBは未刺激の状態では細胞質に存在しており、刺激によって核に移行することで活性化されます。未刺激時にはNF-κBはIκB (inhibitor of κB)ファミリーメンバーのIκBαなどの阻害タンパク質と結合して細胞質に存在しています。細胞がTNF-α、IL-1β、リポポリサッカライド(LPS)などToll-like受容体と結合するリガンドによって刺激されるとIKK(IκB kinase)複合体が活性化され、阻害タンパク質(IκBαなど)をリン酸化します。その結果、IκBαが分解されることで、NF-κBが阻害タンパク質から遊離し、阻害タンパク質によって隠されていた核移行シグナルが表在化して、NF-κBが核に移行して活性化されます(図1)。刺激依存的にリン酸化されたIκBαのユビキチン化、プロテアソームによる分解、NF-κBの核移行に関してはほぼ定説として確立されています。
刺激依存的なIκBのリン酸化を司るのはIKK1(IKKα)、IKK2 (IKKβ)、NEMO(IKKγ)の3種のタンパク質から構成されているIKK複合体です。IKK1、IKK2はリン酸化酵素ですが、IκBをリン酸化するのはIKK2であると考えられています。NEMOはリン酸化酵素としての活性を持っておらずIKK複合体の活性調節因子であると考えられています。種々の刺激よってIKK複合体が活性化されるメカニズムに関してはまだ、コンセンサスとなっている説はありませんでした。私たちはLUBACが刺激依存的にNEMOを直鎖状ポリユビキチン化することでIKK複合体が活性化され、NF-κBの活性化を引き起こすことを提唱しています。
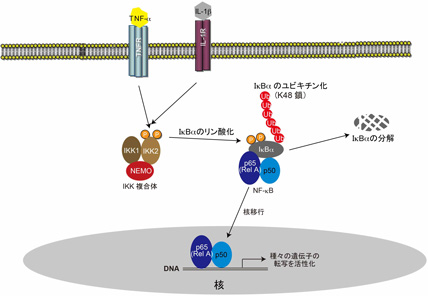
図1 : 刺激依存的NF-κB活性化機構
a) 直鎖状ポリユビキチン鎖とは
まず、私たちが見出した直鎖状ポリユビキチン鎖について簡単に紹介します。ユビキチン修飾系の説明でも紹介しているように、ユビキチン修飾系の大きな特徴はユビキチンが数珠状に連なったユビキチンのポリマーであるポリユビキチン鎖を結合することで標的タンパク質の機能を制御する場合が多い点にあり、ポリユビキチン鎖の種類によってタンパク質の機能制御様式が異なると考えられています。
これまで知られていたポリユビキチン鎖はユビキチンのリシン側鎖にユビキチンのC末端がイソペプチド結合して形成されます。私たちが新たに同定したLUBACユビキチンリガーゼ複合体がユビキチンのN末端のメチオニンのα-アミノ基にユビキチンのC末端をペプチド結合させて、直鎖状ポリユビキチン鎖を形成することを明らかにしました。
b) 直鎖状ポリユビキチン鎖によるNF-κB活性化機構
直鎖状ポリユビキチン鎖を選択的に生成するLUBACユビキチンリガーゼはHOIL-1L、HOIP、SHARPINから構成されています。私たちはLUBACの構成成分であるHOIL-1Lノックアウトマウスなどを用いて、LUBACはTNF-αなどの刺激依存的にIKK複合体のNEMOを識別して直鎖状ポリユビキチン化することがNF-κBの活性化に関与することを提唱しています。(図2)
また、SHARPINは慢性皮膚炎や、種々の免疫異常を示し、自己炎症症候群様症状と免疫不全を呈する自然変異マウスであるcpdmマウス(図3)の責任遺伝子です。SHARPINが欠損することでLUBACユビキチンリガーゼが減少することが、cpdmマウスの表現系を引き起こす事が判っています。言い換えれば、おそらく直鎖状ポリユビキチン鎖形成不全により、個体の恒常性が破綻し、慢性炎症症状が出現すると考えられます。
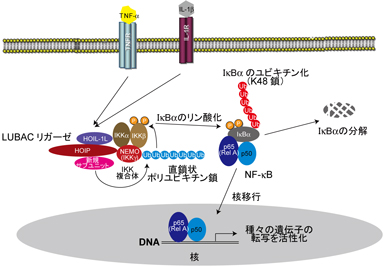
図2 : 直鎖状ポリユビキチン鎖NF-κB活性化における役割

図3 : cpdmマウスの皮膚症状