
実はユビキチン修飾の生化学ではa. ポリユビキチン鎖の形成機構とb. 形成されるポリユビキチン鎖の種類の決定様式は未だに謎になっています(図5)。
LUBACは以下の2つの性質を有しています。
1. LUBACはリジン側鎖を介したポリユビキチン鎖は形成せず、直鎖状ポリユビキチン鎖のみを形成できること。
2. LUBACはユビキチンを識別してその識別したユビキチンのN末端にユビキチンを付加することができること。
したがって、LUBACをモデル系に用いることで、前述の2つの問題にアプローチすることが出来るので、すでに樹立している直鎖状ポリユビキチン鎖の試験管内生成系を用いて解析を進めています。
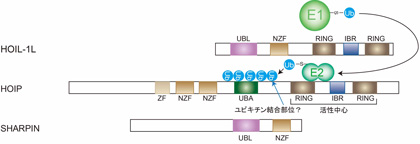
図5 : LUBACによる直鎖状ポリユビキチン鎖伸長メカニズム
LUBACはHOIL-1L、HOIP、SHARPINから構成されます。LUBACはE3(ユビキチンリガーゼ)ですが、他のユビキチンリガーゼとは異なり、標的タンパク質を選択的に認識するだけではなく、生成するポリユビキチン鎖の種類も決定します。LUBACの活性中心サブユニットであるHOIPがユビキチンを認識して直鎖状ポリユビキチン鎖を生成する。
- 私たちがこれまで進めてきた研究の概略についてです。
a. ユビキチン修飾系による選択的な基質識別メカニズム
先にも述べましたが、ユビキチンリガーゼによる時を得た選択的な基質識別はユビキチン修飾系が多彩な機能を発現できる大きな理由の1つです。ユビキチンリガーゼによる標的タンパク質の選択的識別にはリン酸化などの標的タンパク質の翻訳後修飾がシグナルとなることが知られています。私たちは
1.鉄代謝制御因子IRP2の鉄依存性分解の研究を進め、IRP2がヘムと酸素によって受ける酸化修飾が鉄依存的なシグナルとして機能することを示して来ました。
2.東京都医学総合研究所の吉田雪子博士と共同で、タンパク質に付加されたN型糖鎖も選択的なユビキチン化のシグナルとなることを明らかにしてきました。N型糖鎖はERで付加され、膜コンパートメントに存在するタンパク質、分泌タンパク質のほとんどはN型糖鎖を持っています。ユビキチン修飾系は細胞質、核に存在するのでN型糖鎖とは存在部位が異なり原則的には出会うことがありません。ですからN型糖鎖が選択的なユビキチン修飾のシグナルになり得るのです。
b. ユビキチン修飾系の異常によるガン、神経変性疾患の発症メカニズム
ユビキチンリガーゼと疾患について進めてきた研究を紹介しましょう。
1. ユビキチンリガーゼ異常による発ガン
von Hippel-Lindau(VHL)病は腎細胞癌、褐色細胞腫、網膜の血管腫、中枢神経系の血管芽腫など種々の良性悪性腫瘍により特徴づけられる常染色体優性遺伝性疾患です。その原因遺伝子として同定されたVHLは80%以上の散発性の腎淡明細胞癌や小脳の血管芽腫においても欠損または発現抑制が報告されている癌抑制性遺伝子です。VHL遺伝子の産物pVHLはelonginB/C, Cullin2, Rbx1とVBC-Cul2複合体を形成することが知られていました。私たちはVBC-Cul2複合体がユビキチンリガーゼ活性を有しており,pVHLはその基質認識サブユニットとして機能していることを示し、ユビキチンシステムの分子の異常が発ガンの引き金を弾くとの知見を初めて明らかにしました(図6)。
さらに、VBC-Cul2リガーゼが低酸素応答性転写因子であるHIF-αを酸素依存性にユビキチン修飾するリガーゼであることを示し,pVHL欠損により、細胞が正常酸素状態でもあたかも低酸素であるかのように感じることがガン化に引き金を引くことを明らかにしました。VBC-Cul2リガーゼはプロリン残基がヒドロキシル化されたHIF-αを選択的に識別されることが報告されています。